Akonitase
| Akonitase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
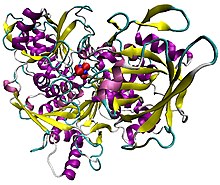 Ilustrasi akonitase khinzir sebagai kompleks dengan kluster [Fe4S4]. Protein ini diwarnakan mengikut struktur sekunder, dengan atom besi berwarna biru, dan atom sulfur berwarna merah.[1] | |||||||||
| Pengenal pasti | |||||||||
| Nombor EC | 4.2.1.3 | ||||||||
| Nombor CAS | 9024-25-3 | ||||||||
| Pangkalan data | |||||||||
| IntEnz | Lihat IntEnz | ||||||||
| BRENDA | Entri BRENDA | ||||||||
| ExPASy | Lihat NiceZyme | ||||||||
| KEGG | Entri KEGG | ||||||||
| MetaCyc | Laluan metabolik | ||||||||
| PRIAM | Profil | ||||||||
| Struktur PDB | RCSB PDB PDBj PDBe PDBsum | ||||||||
| Ontologi gen | AmiGO / EGO | ||||||||
| |||||||||
| Keluarga akonitase (akonitat hidratase) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Struktur akonitase[2] | |||||||||
| Pengenal pasti | |||||||||
| Simbol | Aconitase | ||||||||
| Pfam | PF00330 | ||||||||
| InterPro | IPR001030 | ||||||||
| PROSITE | PDOC00423 | ||||||||
| SCOP | 1aco | ||||||||
| SUPERFAMILY | 1aco | ||||||||
| |||||||||
Akonitase (akonitat hidratase; EC 4.2.1.3) ialah enzim yang memangkinkan pengisomeran stereokhusus sitrat kepada isositrat melalui cis-akonitat dalam kitaran asid trikarboksilik, sebuah proses aktif bukan redoks.[3][4][5]
-
 Asid sitrik
Asid sitrik -
 Asid akonitik
Asid akonitik -
 Asid isositrik
Asid isositrik



Struktur
Akonitase seperti yang dipaparkan dalam struktur di belah kanan halaman ini, mempunyai dua struktur yang sedikit berbeza, bergantung pada sama ada ia diaktifkan atau dinyahaktifkan.[6][7] Dalam bentuk tidak aktif, strukturnya dibahagikan kepada empat domain.[6] Mengira dari terminal N, hanya tiga domain pertama ini terlibat dalam interaksi rapat dengan kelompok [3Fe-4S], tetapi tapak aktif terdiri daripada sisa daripada keempat-empat domain, termasuk domain terminal C yang lebih besar.[6] Kelompok Fe-S dan anion SO2−
4 juga berada di tapak aktif.[6] Apabila enzim diaktifkan, ia memperoleh atom besi tambahan, mewujudkan gugusan [4Fe-4S].[7][8] Walau bagaimanapun, struktur enzim yang lain hampir tidak berubah; atom terpelihara antara kedua-dua bentuk pada dasarnya berada pada kedudukan yang sama, sehingga perbezaan 0.1 angstrom.[7]
Fungsi
Berbeza dengan kebanyakan protein besi-sulfur yang berfungsi sebagai pembawa elektron, kelompok besi-sulfur akonitase bertindak balas secara langsung dengan substrat enzim. Akonitase mempunyai gugusan [Fe4S4]2+ aktif, yang boleh bertukar kepada bentuk [Fe3S4]+ yang tidak aktif. Tiga sisa sisteina (Cys) telah ditunjukkan sebagai ligan pusat [Fe4S4]. Dalam keadaan aktif, ion besi labil gugusan [Fe4S4] tidak diselaraskan oleh Cys tetapi oleh molekul air.
Protein pengikat unsur responsif besi (IRE-BP) dan 3-isopropilmalat dehidratase (α-isopropilmalat isomerase; EC 4.2.1.33), enzim yang memangkinkan langkah kedua dalam biosintesis leusina, dikenali sebagai homolog akonitase. Unsur kawal atur besi (IRE) membentuk keluarga struktur gelung batang 28 nukleotida bukan pengekod yang mengawal penyimpanan besi, sintesis hem dan pengambilan besi. Ia juga mengambil bahagian dalam pengikatan ribosom dan mengawal perolehan mRNA (penguaraian). Protein kawal atur khusus, IRE-BP, mengikat kepada IRE di kedua-dua kawasan 5' dan 3', tetapi hanya kepada RNA dalam bentuk apo, iaitu tanpa kelompok Fe-S. Ekspresi IRE-BP dalam sel kultur telah mendedahkan bahawa protein berfungsi sama ada sebagai akonitase aktif apabila sel penuh dengan zat besi, atau sebagai protein pengikat RNA aktif apabila sel kekurangan zat besi. Mutan IRE-BP, di mana mana-mana atau kesemua tiga sisa Cys yang terlibat dalam pembentukan Fe-S digantikan oleh serina, tidak mempunyai aktiviti akonitase, tetapi mengekalkan sifat mengikat RNA.
Akonitase dihalang oleh fluoroasetat, menyebabkan fluoroasetat beracun. Fluoroasetat ditukar kepada fluorositrat oleh sitrat sintase dalam kitaran asid sitrik. Fluorositrat secara kompetitif menghalang akonitase lalu menghentikan kitaran asid sitrik.[9] Kelompok sulfur besi sangat sensitif terhadap pengoksidaan oleh superoksida.[10]
Mekanisme

Akonitase menggunakan mekanisme penyahhidratan-penghidratan.[11] Sisa pemangkin yang terlibat ialah His-101 dan Ser-642.[11] His-101 memprotonkan kumpulan hidroksil dalam C3 sitrat, membenarkan ia keluar sebagai air, dan Ser-642 serentak mengabstrakkan proton C2, mewujudkan ikatan berganda antara C2 dan C3, dan membentuk apa yang dipanggil sebagai perantara cis-akonitat (dua kumpulan karboksil pada ikatan ganda dua ialah cis ).[11][14] Atom karbon dari mana hidrogen dikeluarkan adalah yang berasal dari oksaloasetat dalam langkah sebelumnya dalam kitaran asid sitrik, bukan yang berasal dari asetil KoA, walaupun kedua-dua karbon ini setara kecuali yang satu adalah "pro-R" dan yang lain pula adalah "pro-S".[15]:393 Pada ketika ini, perantara diputar 180°.[11] Putaran ini dirujuk sebagai satu "penerbalikan".[12] Disebabkan penerbalikan ini, perantara dikatakan bergerak daripada "mod sitrat" kepada "mod isositrat."[16]
Bagaimana sebenarnya penerbalikan ini berlaku dapat dipertikaikan. Satu teori ialah, dalam langkah pengehad kadar mekanisme, cis-akonitat dibebaskan daripada enzim lalu dicantumkan semula dalam mod isositrat untuk melengkapkan tindak balas.[16] Langkah pengehad kadar ini memastikan bahawa stereokimia yang betul, khususnya (2R,3S), terbentuk sebagai produk akhir.[16][17] Satu hipotesis lain ialah cis-akonitate kekal terikat di enzim semasa ia terbalik daripada sitrat kepada mod isositrat.[11]
Dalam kedua-dua kes, penerbalikan cis-akonitat membolehkan langkah penyahhidratan dan penghidratan berlaku di muka yang bertentangan dengan perantara.[11] Akonitase memangkinkan penyingkiran/penambahan trans air, dan penerbalikan menjamin bahawa stereokimia yang betul terbentuk dalam produk.[11][12] Untuk melengkapkan tindak balas, sisa serina dan histidina membalikkan tindakan pemangkin asalnya: histidina, kini sebagai bes, mengabstrak proton daripada air lalu menyediakannya sebagai nukleofil untuk menyerang C2, dan serina terproton dinyahproton oleh ikatan ganda dua cis-akonitat untuk melengkapkan penghidratan, menghasilkan isositrat.[11]

Ahli keluarga
Akonitase diekspresikan dalam bakteria kepada manusia. Manusia menyatakan dua isozim akonitase berikut:
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Rujukan
- ^ PDB: 7ACN; Lauble, H.; Kennedy, M. C.; Beinert, H.; Stout, C. D. (1992). "Crystal structures of aconitase with isocitrate and nitroisocitrate bound". Biochemistry. 31 (10): 2735–48. doi:10.1021/bi00125a014. PMID 1547214.
- ^ PDB: 1ACO; Lauble, H; Kennedy, MC; Beinert, H; Stout, CD (1994). "Crystal Structures of Aconitase with Trans-aconitate and Nitrocitrate Bound". Journal of Molecular Biology. 237 (4): 437–51. doi:10.1006/jmbi.1994.1246. PMID 8151704.
- ^ "Aconitase, a two-faced protein: enzyme and iron regulatory factor". FASEB Journal. 7 (15): 1442–9. Dec 1993. doi:10.1096/fasebj.7.15.8262329. PMID 8262329.
- ^ Flint, Dennis H.; Allen, Ronda M. (1996). "Iron−Sulfur Proteins with Nonredox Functions". Chemical Reviews. 96 (7): 2315–34. doi:10.1021/cr950041r. PMID 11848829.
- ^ "Aconitase as Ironminus signSulfur Protein, Enzyme, and Iron-Regulatory Protein". Chemical Reviews. 96 (7): 2335–2374. Nov 1996. doi:10.1021/cr950040z. PMID 11848830.
- ^ a b c d "The structure of aconitase". Proteins. 5 (4): 289–312. 1989. doi:10.1002/prot.340050406. PMID 2798408.
- ^ a b c "Structure of activated aconitase: formation of the [4Fe-4S] cluster in the crystal". Proceedings of the National Academy of Sciences of the United States of America. 86 (10): 3639–43. May 1989. Bibcode:1989PNAS...86.3639R. doi:10.1073/pnas.86.10.3639. PMC 287193. PMID 2726740.
- ^ "Crystal structures of aconitase with isocitrate and nitroisocitrate bound". Biochemistry. 31 (10): 2735–48. Mar 1992. doi:10.1021/bi00125a014. PMID 1547214.
- ^ "Biochemistry of fluoroacetate poisoning: the effect of fluorocitrate on purified aconitase". Biochem. J. 58 (3): 473–9. November 1954. doi:10.1042/bj0580473. PMC 1269923. PMID 13208639.
- ^ Gardner, Paul R. (2002). "Aconitase: Sensitive target and measure of superoxide". Superoxide Dismutase. Methods in Enzymology. 349. m/s. 9–23. doi:10.1016/S0076-6879(02)49317-2. ISBN 978-0-12-182252-1. PMID 11912933.
- ^ a b c d e f g h i Takusagawa F. "Chapter 16: Citric Acid Cycle" (PDF). Takusagawa’s Note. The University of Kansas. Diarkibkan daripada yang asal (PDF) pada 2012-03-24. Dicapai pada 2011-07-10.
- ^ a b c "Aconitase as Ironminus signSulfur Protein, Enzyme, and Iron-Regulatory Protein" (PDF). Chemical Reviews. 96 (7): 2335–2374. Nov 1996. doi:10.1021/cr950040z. PMID 11848830. Diarkibkan daripada yang asal (PDF) pada 2011-08-11. Dicapai pada 2011-05-16.
- ^ a b PDB: 1C96; "The mechanism of aconitase: 1.8 A resolution crystal structure of the S642a:citrate complex". Protein Sci. 8 (12): 2655–62. December 1999. doi:10.1110/ps.8.12.2655. PMC 2144235. PMID 10631981.
- ^ "Sites and mechanisms of aconitase inactivation by peroxynitrite: modulation by citrate and glutathione". Biochemistry. 44 (36): 11986–96. Sep 2005. doi:10.1021/bi0509393. PMID 16142896.
- ^ Lubert Stryer (1981). Biochemistry (ed. 2nd). m/s. 295–296.
- ^ a b c "Steric and conformational features of the aconitase mechanism". Proteins. 22 (1): 1–11. May 1995. doi:10.1002/prot.340220102. PMID 7675781.
- ^ "Aconitase family". The Prosthetic groups and Metal Ions in Protein Active Sites Database Version 2.0. The University of Leeds. 1999-02-02. Diarkibkan daripada yang asal pada 2011-06-08. Dicapai pada 2011-07-10.
Pautan luar
- Aconitase dalam Tajuk Subjek Perubatan (MeSH) di Perpustakaan Perubatan Negara AS







